The following guide will attempt to explain what exactly is meant by the term “biomechanical efficiency”. Since this is a complex topic that requires far more space than is available here, the guide will be a simplified explanation, broken down into three parts: part one will introduce the evolution of biomechanics in human ancestors, part two will explore mammalian biomechanics (and how mammals came to dominate most ecosystems), and part three will examine how modern human biomechanics have diverged from those of our ancestors.
Part One
In its simplest definition, biomechanical efficiency is a measure of blood-flow– areas that are efficient receive and release blood very quickly on demand, whereas inefficient areas show interrupted or stymied flow. By a slightly more expanded definition, efficiency is also a measure of musculoskeletal development, lymphatic flow, and muscle response time. However, what truly determines biomechanical efficiency– and is the biomechanical mesh underlying each of these measures— is the human fascial system. The fascial system feeds and drains blood/ lymph (and thus supplies oxygen/ removes waste), builds tissues, and provides its own muscular action. As such, a measure of biomechanical efficiency is actually a measure of fascial development.
Measuring biomechanical efficiency can be very useful for analyzing, understanding, and predicting the patterns of athletes (particularly NFL players). The best players show high levels of efficiency in fascial areas most relevant to their playing position, while poor efficiency can be predictive of non-contact injuries and/ or ineffective traits.
But in order to understand the human fascial system, we first need to understand its evolution. Beginning with our very first vertebrate ancestors and the common ancestor to all non-insect land animals:
Fish
The basic body plan for all amphibians, reptiles, birds, and mammals begins with our common ancestor- fish. Specifically lobe-finned fish (seen in the diagram below).
If you’ve eaten fish, you’re probably at least somewhat familiar with their musculature. All muscle tissue is directly connected to the spine (which is why you’ve got to watch out for bones- usually rib bones- when you’re eating a whole fish). Their method of locomotion is driven by their spinal muscles- one side of their body tenses its muscles while the other side relaxes, then the other side tenses etc. This alternation drives their caudal fin (rear fin) back and forth, propelling them forward in the water. The muscles are directly connected to/ driven by their spine, and by working in alternation, they propel the fish forward.
Tetrapods
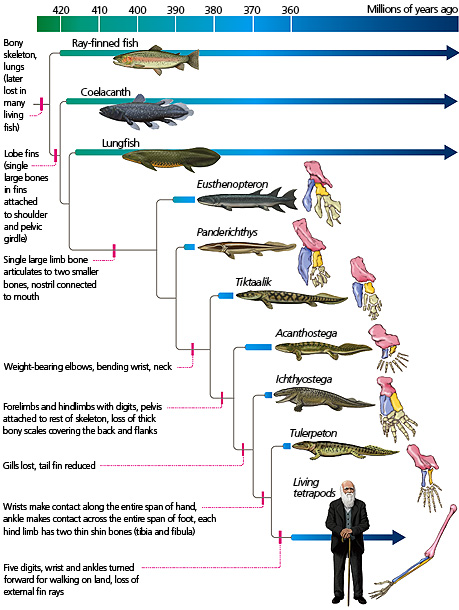
All living land vertebrates- whether they stand on two feet, four feet, or no feet- are considered ‘tetrapods’. This is because our earliest ancestors emerged from the water on four legs. Some eventually returned to the water, some took to the air; some stand on two legs, some on no legs; but all share the same basic body plan- five spinal areas, with all existing limbs connecting via muscle and bone back to these areas.
Amphibians
Amphibians were the first class of tetrapod to live on land, and the first of the extant animal classes to show the basic tetrapod body form- four limbs, a head, a neck, and a tail, all connected to one of five spinal areas . Although today’s amphibians are relatively scarce and occupy few ecological niches, from the Devonian through the early Permian periods, amphibians were the dominant form of life on land.
Biomechanically speaking, amphibians show distinct limb musculature, separate from the spinal muscles. Unlike fish, their limbs are able to be driven via these distinct muscles. However, all motion is still directly linked to the spine via tendons. The spinal muscles still work in alternation, while the limb muscles engage as a direct result of this spinal motion. Because the spine is sectioned into five areas, each of the five portions of the spine are able to fire independently- the tail is driven entirely by the coccygeal spinal muscles, while the head may be raised or lowered at will by the action of the cervical spinal muscles. The thoracic/ lumbar spinal muscles control the movement of the limbs. However, all movement of the limbs/ appendages is still linked directly (via tendons) to the spine.
Consider the characteristic amphibian/ reptilian gait (seen in the video below)- the right foreleg moves with the left hindleg, while the left foreleg moves with the right hindleg. This is the case whether the animal is walking or running- there is no flexibility or ability to adjust this gait, because every linkage is a direct linkage- spinal muscles connect to tendons which connect to limbs.
(notice the alternating fish-like movement of the spine)
Reptiles
Because amphibian skin is permeable and their eggs must be laid in water, they were eventually largely displaced by reptiles (who evolved dry non-permeable skin and hard-shelled eggs that are able to be laid on land). Extant reptiles also show more developed limb musculature than amphibians. However reptiles, like amphibians, derive the entirety of their limb motions from the action of their spinal muscles. The limbs are simply one step removed from the spine via tendons, with spinal action supplemented by limb muscles. There is no ability for independent limb action, separate from the spinal action.
Birds
Birds are often now ordered within the class ‘Reptilia’, even though they were historically considered their own class. And while extant birds are warm-blooded (unlike extant reptiles), with an outer layer of feathers covering a layer of skin and fat (again unlike extant reptiles who instead store fat in their tails), in biomechanical terms, birds are very much like amphibians and reptiles. While the limbs have their own musculature (connected via tendons to the spine), all movement is still derived entirely from the action of the spinal muscles.
Consider the meat of a chicken (or duck, goose, quail, etc). There is a layer of fat directly beneath the skin. But is there any marbling within the muscle? Is there any silverskin to remove- like there is with lamb, beef, or pork? Is there any gristle?
None of the above are found within the meat of birds (or amphibians or reptiles) because the biomechanical system is relatively simple and relatively rigid- all motion is directly tied to one of five spinal areas by tendons, and all spinal action is directly transmitted to the desired limb. As such, all amphibians, reptiles, and birds can be said to be fully efficient by default. There is no biomechanical intermediary between the spinal muscles and the limbs (other than tendons which are essentially direct connections), so barring injury, all spinal action is transmitted fully efficiently to the desired limb/ appendage.
Mammals
So why are marbling, silverskin, and gristle are all to be found in the meat of mammals where are they are absent in the meat of other classes of tetrapods? How did the smooth, light muscles of fish evolve into the dense, chewy, fibrous tissues of mammals? Through a primary evolution of mammals, the fascia.
Fascia are a 3D matrix of tissue surrounding and encasing mammalian muscles and organs. This matrix exists on three levels, with all levels connected to each other via skin ligaments, forming a grid in/ around muscles, organs, and skin. To quote the original primer I wrote in 2016:
“Fascia (I use the term in the plural) form the network of tissue that surrounds and encases internal organs, muscles, and skin. They exist on 3 planes- the superficial, the deep, and the internal. The superficial layers’ musculature is autonomously controlled [(picture goosebumps, or a scared cat)]. Deep fascia show both voluntary and involuntary action and are directly connected to striated muscle tissue.”
This matrix is dynamic- it responds to injury by interjecting itself to protect an injured area. It is also dynamic in that it grows and changes shape in response to behavior and conditions. While two geese from the same flock will move in a virtually identical manner, two identical human twins- sharing the exact same DNA- often move very differently (consider Ronde and Tiki Barber for example). This is because the fascia are constantly shifting in response to changing behaviors/ conditions.
Fascia also show the ability to act as an intermediary between the spinal muscles and limb muscles. This can happen voluntarily- such as when the deep fascia engage and add their own action to the pattern of muscles firing. Or it can happen involuntarily, such as when fascial networks form (in response to trauma or incomplete development), and shape the action of underlying and peripherally-connected muscles.
Fascia act as a dynamic encompassing bridge between the action of spinal muscles and the action of limbs/ appendages. This makes mammalian motion far more changeable and unpredictable. It also makes mammalian motion susceptible to losses of efficiency.
(click here to continue to part 2)